Botanicargentina.com.ar
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff
et al. - Análisis citogeográfico en
Turnera krapovickasii
Bol. Soc. Argent. Bot. 51 (1): 153-167. 2016
AnálIsIs cItogeográfIco en turnerA krApovIckAsII
YANINA LAZAROFF1, E. M. SARA MORENO1,2, AVELIANO FERNÁNDEZ1 y
VIVIANA G. SOLÍS NEFFA1,2
Summary: Cytogeographic analysis in
Turnera krapovikasii (Passifloraceae).
Turnera krapovickasii is
distributed at northwestern Argentina, southern Bolivia and western Paraguay. It presents diploid (2n=
2x= 10) and autotetraploid (2n= 4x= 20) cytotypes. Aiming to interpret the role of autopolyploidy in the
evolution of this species, the frequency and spatial distribution of the cytotypes were analyzed. Equal
frequencies of diploid and tetraploid populations were detected; most of them are constituted by only
one cytotype, although some diploid populations presented triploid or tetraploid individuals. Diploids
occupy a continuous area from southern Bolivia to northwestern Argentina, and from 124 to 1342
m.a.s.l. Tetraploids occur in disjunct areas, in Eastern and northern species area, in altitudes from 237
up to 810 m.a.s.l. The PCA showed a significant association between cytotype distribution and climate.
Moreover, the ecological niche modelling analysis showed that the southeastern (Chaquenean Province)
and eastern Bolivia (Cerrado Province) are the most probable areas of occurrence of diploids and
tetraploids, respectively. The cytotype distribution observed support the hypothesis that the expansion
of the geographic area associated with polyploidy played an important role in the evolutionary history of
Key words:
Turnera krapovickasii, polyploidy, cytogeography
Resumen:
Turnera krapovickasii se distribuye en el noroeste argentino, sur de Bolivia y oeste de
Paraguay. Presenta citotipos diploide (2n= 2x= 10) y autotetraploide (2n= 4x= 20). A fin de interpretar
el papel de la autopoliploidía en la evolución de esta especie, se analizaron la frecuencia y distribución
espacial de los citotipos. Se detectaron iguales frecuencias de poblaciones diploides y tetraploides,
mayormente constituidas por un solo citotipo, aunque algunas poblaciones diploides presentaron
individuos triploides o tetraploides. Los diploides ocupan un área continua desde el sur de Bolivia hasta
el noroeste de Argentina, y desde 124 hasta 1342 m.s.n.m. Los tetraploides ocupan áreas disyuntas,
en el este y norte del área de la especie, en alturas desde 237 hasta 810 m.s.n.m. El ACP mostró una
asociación significativa entre la distribución de los citotipos y el clima. Asimismo, el análisis de modelado
de nicho ecológico reveló que el sureste (Provincia Chaqueña) y el este de Bolivia (Provincia del
Cerrado) son las áreas de mayor probabilidad de ocurrencia de diploides y tetraploides, respectivamente.
La distribución de los citotipos observada sustenta la hipótesis de que la expansión del área geográfica
asociada con la poliploidía tuvo un papel importante en la historia evolutiva de
T. krapovickasii.
Palabras clave:
Turnera krapovickasii, poliploidía, citogeografía
plantas (Stebbins, 1971; Ramsey & Schemske,
1998; Soltis
et al., 2004; Wendel & Doyle,
La poliploidía constituye uno de los principales 2005). Numerosas investigaciones sugirieron que
mecanismos de evolución y diversificación en la poliploidía puede contribuir a la adquisición
de características morfológicas, genéticas y/o
fisiológicas nuevas (Osborn
et al., 2003; Soltis
1Laboratorio de Citogenética y Evolución Vegetal. Instituto
et al., 2004; Adams & Wendel, 2005a, b; Durand
de Botánica del Nordeste (UNNE-CONICET). C.C. 209,
& Hoberman, 2006; Chen & Ni, 2006; Madlung,
3400, Corrientes, Argentina.
2013), las que conferirían a los poliploides una
Facultad de Ciencias Exactas y Naturales y Agrimensura
mayor capacidad competitiva así como una mayor
tolerancia o amplitud ecológica en comparación
Bol. Soc. Argent. Bot. 51 (1) 2016
con la de sus progenitores diploides (Lewis, 1980; – poliploide (Elías
et al., 2011; Kovalsky & Solís
Ehrendorfer, 1980; Levin, 1983; Udall & Wendel, Neffa, 2012). Teniendo en cuenta la distribución
2006). En consecuencia, los poliploides pueden amplia de los tetraploides y la ocurrencia de
presentar una distribución diferente o marginal en los niveles de ploidía más altos en regiones que
comparación con la de los diploides a lo largo de presentan las condiciones más extremas en el
un gradiente ambiental o climático (Levin, 2002; régimen de precipitaciones, se ha sugerido que
Husband
et al. 2013).
los poliploides habrían sido colonizadores más
La poliploidía es también muy frecuente en el efectivos que los diploides, habiendo desempeñado
género
Turnera L. (Passifloraceae). Este género un importante papel en la expansión del área
cuenta con alrededor de 135 especies americanas y geográfica de
T. sidoides (Solís Neffa & Fernández,
dos africanas, las que fueron agrupadas en 9 series 2001; Solís Neffa
et al., 2004; Elías
et al., 2011)
(Urban, 1883). En América, las especies de este y posiblemente también de otras especies con
género se distribuyen desde el sur de los Estados citotipos diploide y autopoliploide de
Turnera. Sin
Unidos hasta el centro - sur de la Argentina, siendo embargo, dado que la frecuencia y distribución de
el área comprendida en la subregión Chaqueña los citotipos puede variar entre grupos taxonómicos,
propuesta por Morrone (2001), la región de mayor incluso entre aquellos muy afines (Husband
et
diversificación del género (Arbo & Espert, 2009).
al., 2013), es preciso contar con información
Los estudios cromosómicos realizados en 35 citogeográfica de otras especies del género a fin de
especies (Raman & Kesavan, 1964; Barrett, 1978; detectar patrones generales y de probar hipótesis
Barrett & Shore, 1987; Arbo & Fernández, 1983; acerca del papel de la poliploidía durante la historia
Fernández, 1987; Solís Neffa & Fernández, 1993; evolutiva de las especies de
Turnera.
Solís Neffa, 1996; Solís Neffa & Fernández,
Turnera krapovickasii Arbo (x= 5) de la serie
2001) permitieron comprobar que en el género
Turnera, constituye un interesante modelo para
existen 3 números básicos (x= 5, 7 y 13) y que investigar la dinámica evolutiva de los poliploides.
aproximadamente el 59 % de las poblaciones El área de distribución de esta especie está
analizadas hasta el momento son de origen comprendida en una región considerada como uno
poliploide, habiéndose detectado desde tetraploides de los centros más importantes de diversificación
hasta decaploides (Solís Neffa & Fernández, de la flora sudamericana (Gentry, 1982; Gentry &
2000). Asimismo, el análisis de la meiosis de los Dodson, 1987; Moraes & Beck, 1992; Killeen
et
poliploides reveló que hay alo- y autopoliploides,
al., 1993; Navarro & Maldonado, 2002; Young
et
presentando algunas especies citotipos diploide y
al., 2002). Se distribuye en el noroeste argentino,
autopoliploide (Fernández, 1987; Shore, 1991a, sur de Bolivia y oeste de Paraguay (Fig. 1). Se
b; Solís Neffa & Fernández, 2000). Sobre la extiende entre los 16º-21ºS y los 59º-65ºW y
base de estos antecedentes, se ha sugerido que la crece en alturas desde el nivel del mar hasta los
poliploidía habría tenido un papel fundamental en 1310 m.s.n.m. (Arbo, 2005). Desde un punto de
la evolución de las especies de
Turnera (Solís Neffa vista biogeográfico, el área de distribución de
T.
& Fernández, 2000, 2001).
krapovickasii comprende parte de las provincias
Actualmente, las investigaciones que se biogeográficas del Pantanal, Cerrado y Chaqueña
desarrollan en
Turnera tienen como uno de sus propuestas por Morrone (2001). Los estudios
objetivos interpretar los mecanismos involucrados cromosómicos mostraron que las poblaciones de
en la dinámica evolutiva y en la diferenciación esta especie, analizadas hasta el momento, son
ecológica de los poliploides. En este marco, se diploides, 2n= 2x= 10, y tetraploides, 2n= 4x= 20
analizaron los patrones de distribución de los (Fernández, 1987; Solís Neffa & Fernández, 1993).
citotipos del complejo
Turnera sidoides L. (x= El análisis de la meiosis de los poliploides demostró
7) de la serie
Leiocarpae Urb., a distintas escalas que son autopoliploides (Fernández, 1987). El
espaciales (Solís Neffa & Fernández, 2001; Solís origen autopoliploide de las accesiones tetraploides
Neffa,
et al.,
2004; Elías
et al., 2011). Dichos fue posteriormente sustentado por un análisis
estudios revelaron que, aunque la mayoría de las filogenético empleando secuencias del genoma de
poblaciones están constituidas por un solo citotipo, cloroplastos (López
et al., 2013).
también se encontraron poblaciones mixtas diploide
Estudios preliminares revelaron que los diploides
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
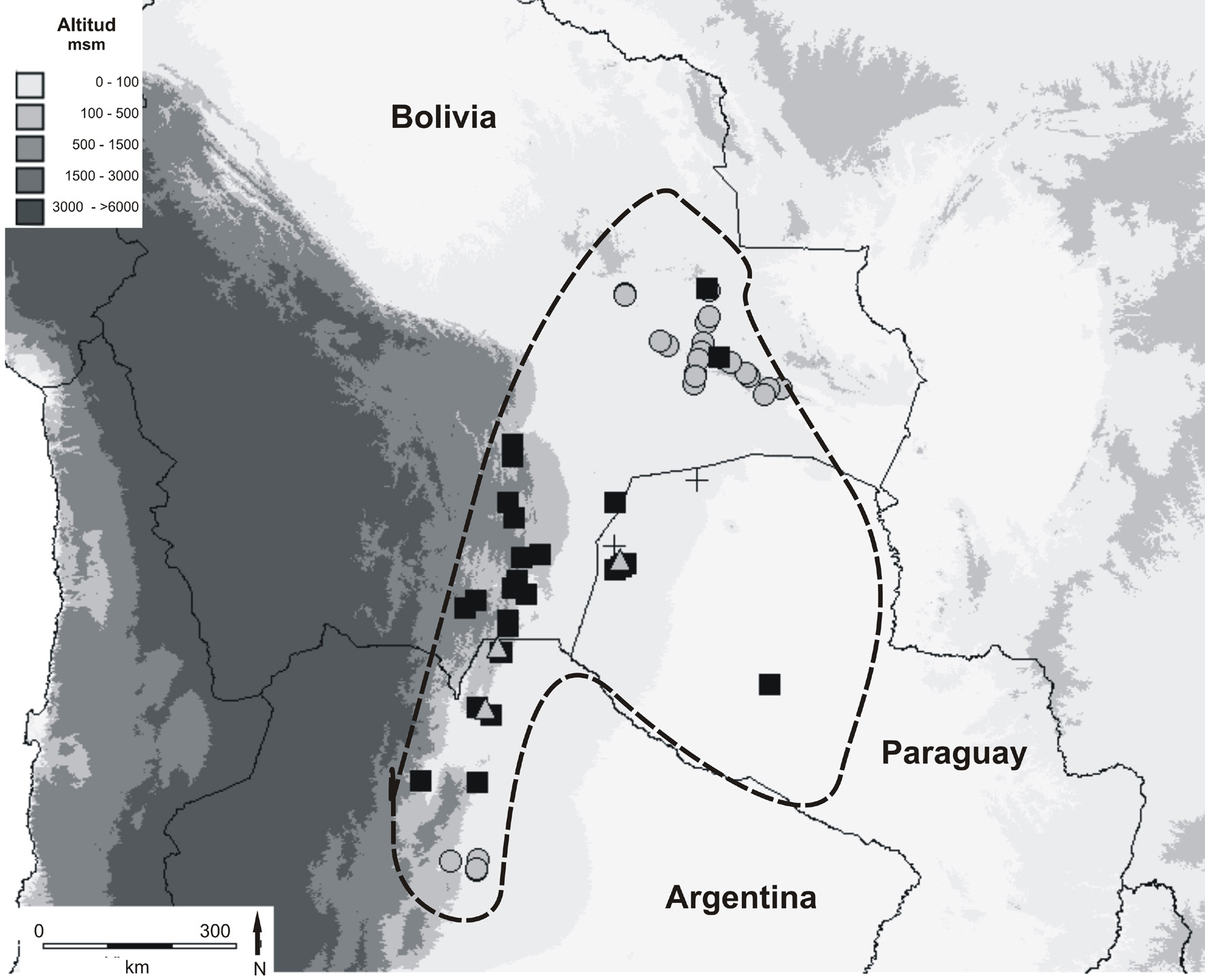
Fig. 1. Principales características geográficas de la zona de distribución de Turnera krapovickasii y
distribución geográfica de los citotipos. La línea punteada indica el área de distribución de T. krapovickasii.
La distribución se basa en muestras de herbario y observaciones de campo. Los distintos citotipos se
indican con figuras: cuadrado = diploide (2n= 2x= 10); círculo = tetraploide (2n= 4x= 20); triángulo =
población mixta diploide - triploide (2n= 3x= 15); cruz = población mixta diploide - tetraploide.
y tetraploides de T. krapovickasii difieren en su
Por lo tanto, a fin de interpretar el papel de
distribución geográfica (Reynoso et al., 2004; la autopoliploidía en la evolución de Turnera
Lazaroff et al., 2009). Dado que el área de esta krapovickasii e inferir los factores que influyen en
especie presenta una gran diversidad de ambientes la distribución de los citotipos de esta especie, en
así como tipos de vegetación relacionados a la este trabajo se analizan la frecuencia y distribución
enorme variedad climática, fisiográfica y altitudinal espacial de los citotipos en relación a variables
(Navarro & Maldonado, 2002), la distribución ambientales.
de los citotipos de T. krapovickasii podría ser el
resultado de diferencias adaptativas de diploides
y tetraploides a la heterogeneidad ambiental. Sin MAterIAles y Métodos
embargo, hasta el momento no se ha realizado un
análisis detallado sobre los factores que determinan
Se analizaron 567 semillas provenientes de
los patrones de distribución de los citotipos en esta 59 poblaciones naturales representativas del área
geográfica de Turnera krapovickasii, obtenidas
Bol. Soc. Argent. Bot. 51 (1) 2016
entre los años 2005 y 2010 (Tabla 1). Algunas (1996) y Fernández & Solís Neffa (2004), se
plantas fueron transportadas a Corrientes representó gráficamente en un mapa confeccionado
(Argentina), donde se cultivaron en el invernáculo con el programa Diva-Gis (Hijmans et al., 2004).
del Instituto de Botánica del Nordeste (IBONE). Los Para evaluar la posible influencia de la altitud y el
ejemplares testigo de los especímenes provenientes clima en la distribución de los citotipos, se registró
de Argentina y Paraguay se encuentran depositados la altitud de cada localidad analizada con un GPS,
en el herbario del citado Instituto (CTES), mientras mientras que las siguientes variables climáticas
que los provenientes de Bolivia fueron depositados fueron extraídas de la base de datos WorldClim
en el Herbario Nacional de Bolivia (LPB). Los (Hijmans et al., 2005): temperatura media anual
duplicados de todos los ejemplares fueron repartidos (BIO 1), rango promedio de temperatura mensual
a otros herbarios del mundo.
(BIO 2), isotermalidad (BIO 3), estacionalidad de
temperatura (BIO 4), temperatura máxima del mes
Recuentos cromosómicos y estimación del nivel de más cálido (BIO 5), temperatura mínima del mes
más frío (BIO 6), rango de temperatura anual (BIO
Se realizaron recuentos cromosómicos en 7), temperatura media del cuatrimestre más húmedo
meiosis en algunos individuos de cada población, (BIO 8), temperatura media del cuatrimestre más
los que fueron utilizados posteriormente como seco (BIO 9), temperatura media del cuatrimestre
patrón para analizar el nivel de ploidía de los más cálido (BIO 10), temperatura media del
restantes individuos de la población por citometría cuatrimestre más frío (BIO 11), precipitación anual
(BIO 12), precipitaciones del mes más húmedo
Los recuentos en meiosis se realizaron a partir de (BIO 13), precipitaciones del mes más seco (BIO
botones florales fijados en alcohol etílico absoluto: 14), estacionalidad de las precipitaciones (BIO 15),
ácido láctico en una proporción 5: 1 (Fernández, precipitaciones del cuatrimestre más húmedo (BIO
1973) y conservados en etanol 70 % a 4 ºC. Los 16), precipitaciones del cuatrimestre más seco (BIO
preparados se obtuvieron por aplastado de las 17), precipitaciones del cuatrimestre más cálido
anteras en carmín acético al 3 %.
(BIO 18) y precipitaciones del cuatrimestre más
La estimación del nivel de ploidía por citometría frío (BIO 19).
de flujo se realizó a partir de semillas a las que
Se realizó un análisis de componentes principales
previamente se extrajo el pericarpio. Tras añadir (ACP) a fin de identificar un número más reducido
0,5 ml de buffer de extracción (Partec UV kit de variables que representen la mayor parte de
CyStain Precise P 05-5002), las muestras fueron la varianza de las variables climáticas entre las
trituradas con una hoja de afeitar. Después de 2 poblaciones. El cálculo del ACP se llevó a cabo
min de incubación, el homogenado fue filtrado a partir de una matriz de correlación estimada
a través de una de malla de nylon de 50 μm de a partir del conjunto de datos estandarizados.
apertura y colocadas en un tubo donde se añadieron Además, para cada citotipo, se calcularon la
1,5 ml de buffer de tinción DAPI (4´,6-diamidino-
media, la desviación estándar promedio y el rango
2-fenilindol). La mezcla fue incubada durante 2 de variación de las variables bioclimáticas. Para
min a temperatura ambiente y luego analizada. evaluar las diferencias entre los citotipos se realizó
Las mediciones de la intensidad de fluorescencia un análisis multivariado de la varianza (MANOVA)
de los núcleos teñidos con DAPI se realizaron con a un nivel de significancia del 5 % (α= 0,05).
un citómetro de flujo Partec PA II (Partec GmbH, Además, para evaluar la existencia de diferencias
Münster, Alemania) con el detector funcionando a significativas entre citotipos para cada una de las
355 nm. Se midieron alrededor de 3000 núcleos por variables, se realizó un análisis de la varianza
muestra. El análisis de los datos se realizó con el (ANOVA) a un nivel de significancia del 5 % (α =
software PA II Partec Flomax.
0,05) luego de realizar la prueba de homogeneidad
de Bartlett. Las diferencias entre cada par de medias
Distribución geográfica de los citotipos
fueron estimadas mediante el test de Tukey al 5
La distribución geográfica de los citotipos %.Todos los análisis estadísticos fueron realizados
hallados en este trabajo junto con los obtenidos utilizando el programa Infostat, versión 2009 (Di
previamente por Fernández (1987), Solís Neffa Rienzo et al., 2009).
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
Tabla 1. Material estudiado de Turnera krapovickasii y sus niveles de ploidía. Los asteriscos (*)
representan estimaciones del nivel de ploidía de nuevas poblaciones. AR: Argentina, BO: Bolivia; PA:
Paraguay. A.
Procedencia y número de colección
Long. O Altitud (m.s.m.) Ploidía
AR, Jujuy, ruta 34, puente sobre Río Negro III. O. Ahumada 1549 (CTES). 24° 05'
AR, Salta, Dpto. San Martín. Campo Durán. A. Panseri, W. Reynoso, M. 22°11'
Sosa & D. Hojsgaard 22 (CTES).
AR, Salta, Río Negro III. A. Panseri, W. Reynoso, M. Sosa & D. Hojsgaard 24°05'
26 (CTES).
AR, Salta, Dpto. San Martín. Ruta 81. A. Panseri, W. Reynoso, M. Sosa & 23°06'
D. Hojsgaard 21 (CTES).
AR, Salta, Dpto. San Martín, camino a Maravilla. A. Panseri, W. Reynoso, 22°11'
M. Sosa & D. Hojsgaard 23 (CTES)AR, Salta. Ruta 81. V. Solís Neffa, G. Seijo, M. Grabiele & W. Reynoso 1975 23º06'
(CTES, ASU, CESJ, ESA, FCQ, HRB, HUEM, JUA, SI).
BO, Dpto. Tarija, O' Connor, 3 km S de Entre Ríos. A. Krapovickas 38858 21° 33' 64° 10'
(CTES).
BO, Dpto. Tarija, Prov. Gran Chaco, Villa Montes. A. Krapovickas 39177 21° 15' 63° 28'
(CTES).
BO, Dpto. Tarija, Prov. O' Connor, 19 km E de Entre Ríos. A. Krapovickas 21° 26' 64° 00'
39099 (CTES).
BO, Dpto. Sta. Cruz, Prov. Cordillera. V. Solís Neffa, G. Seijo, A. Schinini & 19°20'
R. Almada 1062 (CTES).
BO, Dpto. Tarija, Prov. Gran Chaco. V. Solís Neffa & G. Seijo 1482 (CTES, 21° 09'
ASU, ESA, LPB, MA, MO, SI).
BO, Dpto. Tarija, Prov. Gran Chaco. V. Solís Neffa, G. Seijo, M. Grabiele & 21°48'
W. Reynoso 1497 (CTES, LPB, SI, SP).
BO, Dpto. Tarija, Prov. Gran Chaco. V. Solís Neffa, G. Seijo, M. Grabiele & 21°20'
W. Reynoso (CTES, ASU, BAB, ESA, FCQ, GH, HRB, HUEFS, LPB, MBM,
SI).
BO, Dpto. Sta. Cruz, Prov. Cordillera. V. Solís Neffa, G. Seijo, M. Grabiele 19°10'
& W. Reynoso 1935 (CTES, ASU, ESA, FCQ, HRB, HUEM, JUA, LPB, SI,
SP).
BO, Dpto. Sta. Cruz. Prov. Cordillera. Camiri. V. Solís Neffa, G. Seijo, A. 20º00'
Schinini & R. Almada 1055 (CTES, ASU, BA, CANB, LPB, MBM, MICH, SI,
SP).
BO, Dpto. Sta. Cruz, Prov. Cordillera. V. Solís Neffa, G. Seijo, A. Schinini & 20º14'
R. Almada 1432 (CTES, ESA, LPB, SI, SP).
BO, Dpto. Tarija, Prov. Gran Chaco. V. Solís Neffa, G. Seijo, M. Grabiele & 21º43'
W. Reynoso 1503 (CTES, CESJ, FCQ, HRB, LPB, NY, SI, SP).
BO, Dpto. Tarija, Prov. Gran Chaco. V. Solís Neffa, G. Seijo, M. Grabiele & 21º10'
W. Reynoso 1512 (CTES, FCQ, GH, LPB, SI).
BO, Dpto. Chuquisaca, Prov. Azero. V. Solís Neffa, G. Seijo, M. Grabiele & 20º48'
W. Reynoso 1513 (CTES, LPB, MO, NY, SI, SP).
BO, Dpto. Sta. Cruz, Prov. Velasco. V. Solís Neffa, G. Seijo, M. Grabiele & 16º52'
W. Reynoso 1743 (CTES, HUEFS, LPB, SI).
BO, Dpto. Tarija, Prov. Cordillera. V. Solís Neffa, G. Seijo, M. Grabiele & W. 20º46'
Reynoso 1973 (CTES, FCQ, HUEFS, JUA, LPB, MO, SI).
BO, Dpto Tarija. Prov Gran Chaco. V. Solís Neffa s/n.
Bol. Soc. Argent. Bot. 51 (1) 2016
Procedencia y número de colección
Long. O Altitud (m.s.m.) Ploidía
PA, Dpto. Presidente Hayes. A. Krapovickas & C. Cristóbal 44366 (CTES). 22° 40' 59° 44'
PA, Dpto. Boquerón. E. Zardini & Acosta 42205 (CTES).
20° 54' 61° 50'
PA, Dpto. Alto Paraguay, Sierra León. Pérez de Molas & G. Navarro 9077 19° 40' 60° 47'
(CTES).
PA, Dpto. Boquerón, Ruta Transchaco. A. Krapovickas, C. Cristóbal & A. 20° 56' 61° 54'
Schinini 45380 (CTES).
PA, Dpto. Boquerón. A. Krapovickas, C. Cristóbal & A. Schinini 45462 20° 56'
(CTES).
PA, Dpto. Alto Paraguay, Parque Cué. F. Mereles & Degen 5979 (CTES).
20° 07' 61° 41'
AR, Salta. A. Panseri, W. Reynoso, M. Sosa & D. Hojsgaard 25 (CTES).
AR, Salta, Dpto. San Martín. Río Conepoi. A. Panseri, W. Reynoso, M. Sosa 22°06'
& D. Hojsgaard 24 (CTES).
PA, Dpto. Boquerón. Schinini & Palacios 25640 (CTES).
PA, Dpto. Boquerón, Transchaco. A. Krapovickas, C. Cristóbla & A. Schinini 20° 38'
45430 (CTES).
PA, Dpto. Alto Paraguay, Sierra León. Perez de Molas & G. Navarro 9084 19° 40' 60° 47'
(CTES).
AR, Salta, Dpto. Anta. El Quebrachal. V. Solís Neffa, G. Seijo, M. Grabiele & 25º21'
W. Reynoso 1478 (CTES, ESA, MO, SI).
AR, Salta, Dpto. Anta. A. Panseri, W. Reynoso, M. Sosa & D. Hojsgaard s/n. 25°12'
AR, Salta, el Tunal, Río Juramento. A. Panseri, W. Reynoso, M. Sosa & D. 25°14'
Hojsgaard 28 (CTES).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, A. Schinini & 17º42'
R. Almada 1315 (CTES, BAB, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. San José de Chiquitos. V. Solís Neffa, 18° 09' 60° 50'
G. Seijo, A. Schinini & R. Almada 1285 (CTES, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Ñuflo de Chaves. V. Solís Neffa, G. Seijo, M. 16°56'
Grabiele & W. Reynoso 1546 (CTES, HRB, K, LPB, MBM, SI).
BO, Dpto. Sta. Cruz, Prov. Ñuflo de Chaves. V. Solís Neffa, G. Seijo, M. 16°58'
Grabiele & W. Reynoso 1547 (CTES, LPB, SI, ZT).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18°09'
W. Reynoso 1917 (CTES, ASU, FCQ, LPB,MO, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18°06'
W. Reynoso 1922 (CTES, BAB, CESJ, GH, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17°57'
W. Reynoso 1927 (CTES, FCQ, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º52'
W. Reynoso 1876 (CTES, GH, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, A. Schinini & 17º47'
R. Almada 1273 (CTES, BAA, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. San José de Chiquitos. V. Solís Neffa, 17° 54' 60° 47'
G. Seijo, A. Schinini & R. Almada 1278 (CTES, LPB, MO, SI).
BO, Dpto. Sta. Cruz, Prov. Velasco. V. Solís Neffa, G. Seijo, M. Grabiele & 16º54'
W. Reynoso 1749 (CTES, JUA, LPB, NY, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º17'
W. Reynoso 1766 (CTES, LPB, SI, MA).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º40'
W. Reynoso 1770 (CTES, LPB, SI).
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
Procedencia y número de colección
Long. O Altitud (m.s.m.) Ploidía
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º47'
W. Reynoso 1777 (CTES, LPB).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. Serranías de Chiquitos. V. Solís Neffa, 17° 54' 60° 47'
G. Seijo, M. Grabiele & W. Reynoso 1784 (CTES, LPB).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18º07'
W. Reynoso 1796 (CTES, LPB).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18º09'
W. Reynoso 1799 (CTES, LPB, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18º16'
W. Reynoso 1866 (CTES, LPB, SI),BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º52'
W. Reynoso 1877 (CTES, LPB, SI, HUEFS).
BO, Dpto. Santa Cruz. Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele 18º25'
& W. Reynoso 1888 (CTES, LPB, SI)BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 18º19'
W. Reynoso 913 (CTES, LPB)BO, Dpto. Sta. Cruz, Prov. Chiquitos. Roboré. V. Solís Neffa, G. Seijo, M. 18º20'
Grabiele & W. Reynoso 1915 (CTES, LPB, NY, SI).
BO, Dpto. Sta. Cruz, Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele & 17º38'
W. Reynoso 1932 (CTES, ESA, JUA, LPB, NY, SI, SP).
BO, Dpto. Santa Cruz. Prov. José Miguel de Velasco. V. Solís Neffa, G. 17º22'
Seijo, A. Schinini & R. Almada 1266 (CTES, LPB, SI)BO, Dpto. Santa Cruz. Prov. Chiquitos. V. Solís Neffa, G. Seijo, M. Grabiele 18º38'
& W. Reynoso 1908 (CTES, JUA, LPB, SI).
BO, Dpto. Santa Cruz. Prov. Nuflo de Cháves. V. Solís Neffa, G. Seijo, M. 16º54'
Grabiele & W. Reynoso 1545 (CTES, K, LPB, MA, NY, SI).
PA, Dpto. Nueva Asunción. Gral E.A. Garay. S. Beck 9433.
Finalmente, la distribución geográfica potencial solo citotipo, mientras que en otras poblaciones
actual de los citotipos de T. krapovickasii fue se detectó la presencia de individuos triploides
predicha mediante el modelado del nicho ecológico (2n= 3x= 15, 4,55 %) o tetraploides (3,03 %) en
utilizando el programa DIVA-GIS (Hijmans et al. algunas poblaciones diploides. Sin embargo, hasta
el momento, no se encontraron plantas adultas
triploides o tetraploides en dichas poblaciones
diploides. Todas las poblaciones tetraploides
estaban constituidas por un solo citotipo.
Recuentos cromosómicos y estimación del nivel de Distribución geográfica de los citotipos
El análisis de la distribución geográfica de
Los resultados obtenidos en este trabajo los citotipos de T. krapovickasii mostró que las
mostraron que las poblaciones de T. krapovickasii poblaciones diploides están concentradas en el
son diploides (2n= 2x= 10; 50 %) y tetraploides oeste del área de distribución de la especie. Dichas
(2n= 4x= 20; 50 %) (Fig. 2; Tabla 1). Asimismo, poblaciones se extienden desde los 19º S en Bolivia
los resultados de la estimación del nivel de ploidía y se prolongan hacia el sur por el noroeste de
a partir de semillas, reveló que la mayoría de Argentina siguiendo la franja oriental de las Sierras
las poblaciones diploides (42,42 % del total de Subandinas hasta los 24º S, y entre los 59º W y 64º
poblaciones analizadas) están constituidas por un W, en alturas que varían entre los 124 hasta los 1342
Bol. Soc. Argent. Bot. 51 (1) 2016
Fig. 2. Estimación del nivel de ploidía por meiosis y citometría de flujo en Turnera krapovickasii. A, citotipo
dipoide (2n= 2x= 10), diacinesis con 5II. B, citotipo tetraploide (2n= 4x= 20), diacinesis con 5II+2IV. C, D
y E histogramas obtenidos por citometría de flujo a partir de semillas. C, semilla con embrión diploide y
endosperma triploide. D, semilla con embrión triploide y endosperma pentaploide. E, semilla con embrión
tetraploide y endosperma triploide hexaploide. Barra 5 μm.
m.s.n.m. Por otra parte, las poblaciones tetraploides %) y separa a las poblaciones tetraploides de
ocupan áreas disyuntas, en el este y norte del área Bolivia y Paraguay (ubicadas hacia la derecha del
de distribución de la especie, en alturas que varían diagrama), de las diploides y de un grupo más
entre los 237 y 810 m.s.n.m. En el norte de Argentina pequeño de poblaciones tetraploides de Argentina
y oeste de Paraguay se hallaron poblaciones mixtas (ubicadas hacia la izquierda del diagrama). La
2x-3x y 2x-4x, ubicadas en altitudes intermedias, isotermalidad, la temperatura mínima del mes
no superiores a los 511 m.s.n.m. (Fig. 1).
más frío, la temperatura media del cuatrimestre
El MANOVA realizado con las variables más frío, las precipitaciones del mes más seco,
climaticas mostró diferencias significativas entre las precipitaciones del cuatrimestre más seco
los citotipos (λ Wilks = 8,61; p < 0.00001). y las precipitaciones del cuatrimestre más frío
Asimismo, el ANOVA mostró que 15 de las están correlacionadas positivamente con este eje,
20 variables climáticas analizadas, presentaron mientras que la estacionalidad de la temperatura,
diferencias significativas entre citotipos (p<= 0,05, el rango de temperatura anual y la estacionalidad
Tabla 2). El ACP basado en los datos bioclimáticos de la precipitaciones lo están negativamente. El
(Fig. 3) mostró que el primer eje de ordenación segundo eje representa el 28,3 % de la variabilidad
explica la mayor parte de la variación (54,1 de los datos climáticos, y agrupa a la mayoría de
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
Tabla 2. Altitud y variables bioclimáticas analizadas en el área geográfica de los citotipos de Turnera
krapovickasii. Se representa la media, desviación estándar y el rango de variación de cada variable. Ver
referencias en Materiales y Métodos.
F (ANOVA) p
200,51 – 423,51 337,00 – 421,77 295,23 – 332,78 193,37 – 464,64
BIO 12 (mm³) 772,89 ± 193,39 813,67 ± 285,10
465,00 – 1308,00 509,00 – 1074,00 483,00 – 580,00 598,00 – 1316,00
BIO 13 (mm³) 145,18 ± 39,49
117,00 – 226,00
BIO 16 (mm³) 405,18 ± 106,88 424,33 ± 156,05
212,00 – 618,00 249,00 – 548,00 236,00 – 237,00 322,00 – 622,00
BIO 18 (mm³) 369,79 ± 96,09
198,00 – 517,00 210,00 – 531,00 201,00 – 226,00 284,00 – 622,00
577,25 ± 310,96 390,67 ± 113,14
132,00 – 1300,00 310,00 – 520,00 234,00 – 335,00 171,00 – 803,00
Bol. Soc. Argent. Bot. 51 (1) 2016
Fig. 3. Ordenamiento de las poblaciones de Turnera krapovickasii en el plano de los primeros componentes
del análisis de Componentes Principales basado en datos bioclimáticos. Los distintos citotipos se indican
con figuras: cuadrado = diploide (2n= 2x= 10); círculo = tetraploide (2n= 4x= 20); triángulo = población mixta
diploide - triploide (2n= 3x= 15); cruz = población mixta diploide - tetraploide. Ver referencias en Materiales
las poblaciones mixtas, algunas diploides y una dIscusIón
tetraploide. Las variables más correlacionadas con
este eje son la temperatura máxima del mes más Recuentos cromosómicos y estimación del nivel de
cálido, la temperatura media del cuatrimestre más ploidía
húmedo y la temperatura media del cuatrimestre
Los resultados obtenidos en este trabajo
más cálido. La correlación cofenética fue alta confirman los recuentos obtenidos previamente los
(r= 0,974), indicando un buen ajuste entre la que demuestran que Turnera krapovickasii posee
distancia euclideana entre las poblaciones en el citotipos diploide y tetraploide (Fernández, 1987;
diagrama de dispersión y la distancia en el espacio Solís Neffa, 1996; Fernández & Solís Neffa, 2004).
multidimensional original.
Además, los resultados del análisis del nivel de
El análisis de modelado de la distribución ploidía por citometría de flujo evidenciaron que la
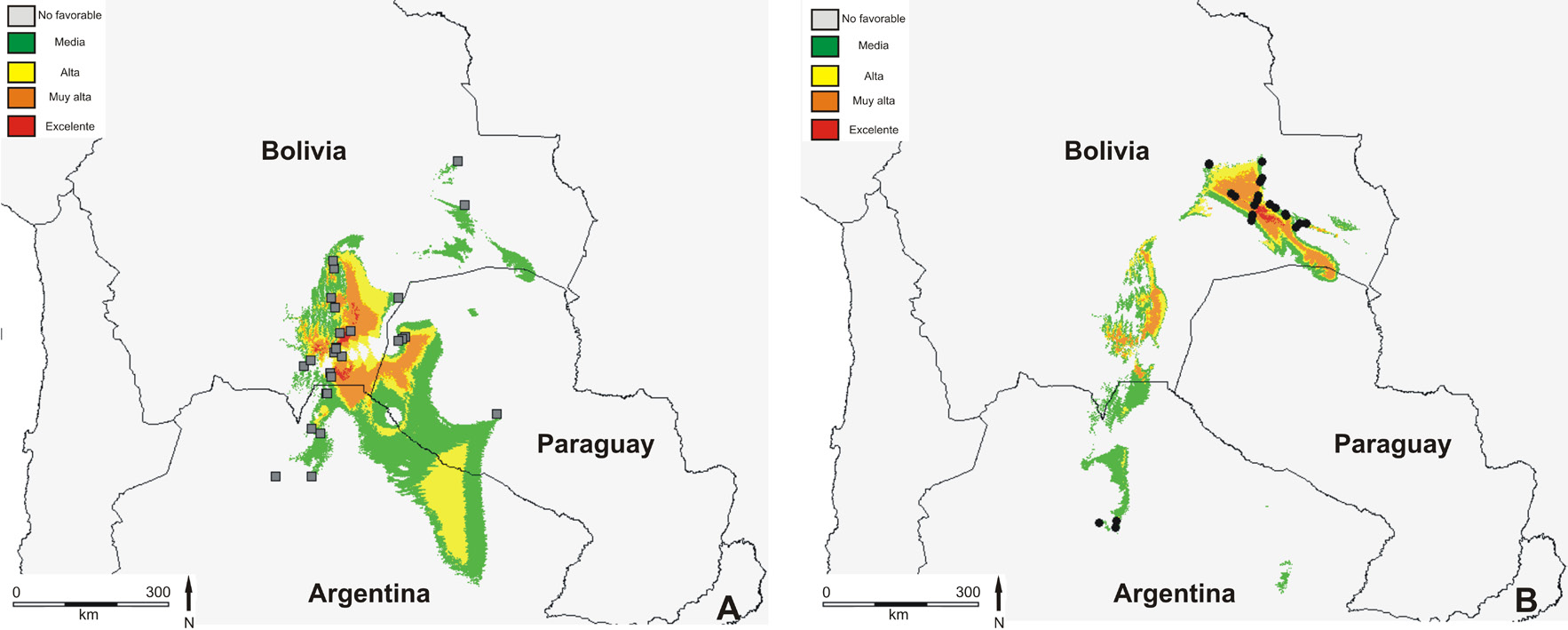
potencial actual mostró que el área de mayor mayoría de las poblaciones de esta especie están
probabilidad de ocurrencia del citotipo diploide constituidas por un sólo citotipo, aunque también
se encuentra al sureste de Bolivia, en la región de se detectaron algunos individuos triploides y
los bosques premontanos (Provincia Biogeográfica tetraploides en las poblaciones diploides. Todos
Chaqueña) que es transicional con las Yungas. A su estos resultados ponen de manifiesto que la
vez, el área de mayor probabilidad de ocurrencia de poliploidía es un fenómeno muy común en T.
los tetraploides se encuentra al este de Bolivia, en la krapovickasii como también lo es en el género
Provincia Biogeográfica del Cerrado, disminuyendo Turnera (Solís Neffa & Fernández, 2000).
esta probabilidad hacia las provincias del Pantanal
Por otra parte, aunque se cuenta con antecedentes
y Chaqueña (Fig. 4).
de la ocurrencia de individuos triploides en
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
Fig. 4. Modelado de la distribución potencial de las poblaciones de Turnera krapovickasii analizadas. A:
Poblaciones diploides. B: Poblaciones tetraploides.
las poblaciones naturales diploides de algunas triploides en las poblaciones diploides podría
especies de Turnera (Elías et al., 2011; Kovalsky deberse a que, como consecuencia de su baja
& Solís Neffa, 2012), los individuos triploides frecuencia, los mismos no fueron detectados
detectados en este trabajo, constituyen el primer en este análisis. Además, también es posible
hallazgo de poliploides impares en poblaciones que los embriones triploides hayan abortado en
naturales de T. krapovickasii. En algunos complejos etapas tempranas del desarrollo como resultado
autopoliploides, los triploides son raros o pueden del desbalance de la relación del nivel de ploidía
estar ausentes (Lumaret et al. 1987; van Dijk et del embrión y del endosperma o el radio materno:
al. 1992), mientras que en otros, pueden aparecer paterno (bloqueo triploide) como fuera propuesto
con mayor frecuencia (Husband & Schemske, para otras especies (Ramsey & Schemske, 1998;
1998). Cuando están presentes, los triploides Köhler et al., 2011). Por otra parte, los tetraploides
pueden formarse como resultado de la unión de hallados en las poblaciones diploides sugieren que
gametos n y 2n dentro de poblaciones diploides la poliploidización sexual bilateral a partir de la
o, por el cruzamiento de diploides y tetraploides unión de dos gametos 2n de los diploides, también
(Ramsey & Schemske, 1998). El hecho que los ocurriría con cierta frecuencia en las poblaciones
triploides de T. krapovickasii fueron detectados en diploides de T. krapovickasii.
las poblaciones diploides sumado a que, en estudios
previos, se detectaron gametos no reducidos en Distribución geográfica de los citotipos
individuos diploides provenientes de las mismas
Los diferentes citotipos de los complejos
poblaciones (Fernández & Arbo, 1990; Lazaroff et poliploides muestran, frecuentemente, marcadas
al., 2011), sustenta el origen de dichos triploides diferencias en su distribución geográfica (Favarger,
por poliploidización sexual unilateral a partir de 1967; Stebbins, 1971; Brunsberg, 1977; Lewis,
la unión de gametos n y 2n de los diploides. Sin 1980; Levin, 1983, 2002; Soltis, 1984; van Dijk et
embargo, aunque en otras especies de Turnera se han al., 1992; Husband & Schemske, 1998; Kessler et
encontrado plantas triploides, tanto en poblaciones al., 2014). En el género Turnera, las investigaciones
naturales como en cruzamientos experimentales llevadas a cabo en el complejo autopoliploide
(Arbo & Fernández, 1987; Fernández & Arbo, T. sidoides demostraron que el citotipo diploide
1990; Elías et al., 2010; Kovalsky & Solís Neffa, posee una distribución más restringida, los
2012), en T. krapovickasii hasta el momento sólo tetraploides poseen una distribución amplia
se encontraron semillas con embriones triploides. ocupando gran parte del área de las subespecies,
El hecho que no se encontraran plantas adultas los hexaploides generalmente están restringidos a
Bol. Soc. Argent. Bot. 51 (1) 2016
regiones marginales, mientras que los octoploides Por otra parte, teniendo en cuenta la abundancia
y poliploides impares son raros (Solís Neffa & relativa de los poliploides en comparación con
Fernández, 2001; Solís Neffa et al., 2004; Elías et sus ancestros diploides es posible estimar la edad
al., 2011). Por otra parte, en T. hassleriana Urban, de un complejo poliploide. Sobre la base de este
T. subulata Smith y T. scabra Millspaugh, los criterio Stebbins (1971) reconoció cinco estados:
citotipos diploide y autotetraploide (Fernández, inicial, joven, maduro, declinante y relictual. De
1987; Shore & Barrett, 1986; Barrett & Shore, 1987; acuerdo con ello, T. krapovickasii y T. scabra
Solís Neffa & Fernández, 1993; Solís Neffa, 1996) serían complejos poliploides jóvenes, en los que
presentan áreas geográficas diferentes, aunque el citotipo diploide está todavía ampliamente
tanto la frecuencia de diploides y tetraploides, distribuido, mientras que los tetraploides se han
como sus patrones de distribución geográfica distribuido más allá del límite de los diploides;
varían entre las especies. En T. hassleriana la única mientras que el complejo T. sidoides y T. subulata
población diploide encontrada hasta el momento serían complejos maduros en los que los diploides
corresponde a una accesión de Bolivia mientras son relictuales y los tetraploides están ampliamente
que las tetraploides son accesiones de Paraguay distribuidos. Estos patrones de distribución
(Fernández, 1987; Solís Neffa & Fernández, 1993). sugieren que el tiempo transcurrido para acumular
En T. subulata, la mayoría de las poblaciones poliploides difiere entre las especies de Turnera.
estudiadas son tetraploides, siendo las poblaciones
Asimismo, diversas hipótesis podrían explicar
diploides y tetraploides simpátricas en parte de las diferencias en la distribución de los citotipos
su área (Fernández, 1987; Shore & Barrett, 1986; diploide y tetraploide de T. krapovickasii. Los
Barrett & Shore, 1987; Solís Neffa & Fernández, primeros estudios sobre el significado de la
1993; Solís Neffa, 1996). Finalmente, en T. scabra variación geográfica de los niveles de ploidía
el 40 % de las poblaciones estudiadas son diploides enfatizaron su relación con la latitud (Hagerup,
y continentales, mientras que el 60 % restante 1932; Tischler, 1935; Stebbins, 1971; Grant,
corresponden a poblaciones insulares tetraploides 1989). Esta relación ha sido interpretada como
(Fernández, 1987; Shore & Barrett, 1986; Barrett una adaptación de los poliploides, en general,
& Shore, 1987). Los resultados de este trabajo a condiciones climáticas más extremas (Lewis,
mostraron que en T. krapovickasii los diploides y 1980). El área de distribución de T. krapovickasii
tetraploides se encuentran en igual frecuencia, las presenta características ecológicas muy diversas
poblaciones diploides en el noroeste de Argentina, debido a su topografía variable y a su ubicación
sureste de Bolivia y oeste de Paraguay, mientras en una zona de transición climática (Killeen
que las tetraploides se encuentran en el este de et al., 1993; Ribera Arismendi, 1993; Navarro
Bolivia y en el noroeste de Argentina. Se ha & Maldonado, 2002). A lo largo del espectro
sugerido que la variación en la frecuencia de latitudinal, esta región presenta un clima tropical
poliploides entre grupos taxonómicos afines sería en el norte y subtropical hacia el sur, variando
el resultado de las diferentes predisposiciones de de húmedo en el este, a semiárido y árido en el
dichos taxones para la evolución por poliploidía oeste y suroeste (Ribera Arismendi, 1993). En T.
o de la diferencia en el tiempo transcurrido krapovickasii, aunque no se observa una tendencia
para acumular poliploides (Meyers & Levin al aumento de la frecuencia de los tetraploides
2006). Ambas hipótesis podrían explicar las con el incremento de la latitud, la distribución
diferencias en la frecuencia de los citotipos entre de los citotipos sería el resultado de diferencias
T. krapovickasii y las otras especies de Turnera adaptativas a las condiciones ambientales ya que
con citotipos diploide y autotetraploide. En primer el ACP basado en los datos bioclimáticos mostró
término, estudios previos en el género Turnera que existe una asociación significativa entre la
demostraron que las especies analizadas producen distribución de los citotipos y las condiciones
diferentes proporciones de gametos no reducidos climáticas de la región. Asimismo, el análisis de
2n y 4n (Fernández et al., 2010; Panseri et al., modelado de la distribución potencial actual reveló
2008; Kovalsky & Solís Neffa, 2012; Lazaroff et que los citotipos poseen diferentes preferencias de
al., inéd.), lo que sugiere que las especies tendrían hábitats. Las poblaciones diploides, que ocupan
diferentes predisposiciones a la poliploidización. un área continua, crecen en lugares de mayor
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
altitud así como más fríos y húmedos en la región BIBlIogrAfíA
de los bosques premontanos de la provincia
Chaqueña. En esta región, las precipitaciones ADAMS, K. L. & J. F. WENDEL. 2005a. Polyploidy and
varían desde 500 mm en el occidente de la planicie
genome evolution in plants: Genome studies and
y en un gradiente creciente hacia el este. La
molecular genetics. Curr. Opin. Pl. Biol. 8: 135–141.
época seca varía desde cuatro hasta ocho meses ADAMS, K. L. & J. F. WENDEL. 2005b. Novel patterns
of gene expression in polyploid plants. Trends
y la temperatura media anual varía desde 22 ºC
Genet. 21: 539–543.
al pie de la Cordillera hasta 26 ºC en el interior ARBO, M. M. 2005. Estudios sistemáticos en Turnera
del Chaco (Ramella & Spichiger, 1989; Killeen
(Turneraceae). III. Series Anomalae y Turnera.
et al., 1993; Navarro & Maldonado, 2002). Los
Bonplandia 14: 115-318.
tetraploides, en cambio, ocupan áreas disyuntas, ARBO, M. M. & A. FERNÁNDEZ. 1983. Posición
aunque la mayoría de las poblaciones se encuentra
taxonómica, citología y palinología de tres niveles
en la provincia biogeográfica del Cerrado cuyo
de ploidía de Turnera subulata Smith. Bonplandia
clima es marcadamente pluviestacional y posee
una época seca de por lo menos tres meses. Las ARBO, M. M. & A. FERNÁNDEZ. 1987. Cruzamientos
intra e interespecíficos en Turnera, serie Canaligerae.
precipitaciones anuales están por encima de 1000
Bonplandia 6: 85-100.
mm al año y la temperatura promedio anual es de ARBO, M. M. & S. M. ESPERT. 2009. Morphology,
alrededor de 22ºC (Ramella & Spichiger, 1989;
phylogeny and biogeography of Turnera
Killeen et al., 1993; Navarro & Maldonado, 2002).
(Turneraceae). Taxon 58: 457-467.
Un número menor de poblaciones tetraploides BARRETT, S. C. H. 1978. Heterostyly in a tropical
fueron encontradas en el sur del área de distribución
weed: the reproductive biology of Turnera ulmifolia
de la especie que presenta un clima más cálido y
complex (Turneraceae). Canad. J. Bot. 1713-1725.
con escasas precipitaciones.
BARRETT, S. C. H. & J. SHORE. 1987. Variation
El hallazgo de poblaciones tetraploides en los
and evolution of breeding systems in the Turnera
ulmifolia L. Complex (Turneraceae). Evolution 41:
extremos norte y sur del área de T. krapovickasii,
sumado a las evidencias citogenéticas y moleculares BRUNSBERG, K. 1977. Biosystematics of the Lathyrus
(Lazaroff et al., ined.), sugiere que los tetraploides
pratensis complex. Opera Bot. Belg. 42: 1–78.
se habrían originado por autopoliploidía de CABRERA, A. L. 1976. Regiones fitogeográficas
manera independiente a partir de poblaciones
argentinas. En: Kugler, W. F. (ed.), Enciclopedia
diploides distribuidas en las áreas marginales de
Argentina de Agricultura y Jardinería, II, ACME,
los diploides. Los tetraploides posteriormente
Buenos Aires, pp. 1-85.
habrían expandido sus áreas hasta alcanzar los CABRERA, A. L. & A. WILLINK. 1973. Biogeografía
limites actuales de T. krapovickasii.
de América Latina. Monografía Nº 13. Secretaría
General de la Organización de los Estados
El patrón de distribución de los citotipos
Americanos. Programa Regional de Desarrollo
detectado en Turnera krapovickasii sustenta la
Científico y Tecnológico, Washington, D.C. pp. 120.
hipótesis de que el rango de expansión asociado CHEN, Z. J. & Z. NI. 2006. Mechanisms of genomic
con la poliploidía juega un papel importante en la
rearrangements and gene expression changes in
historia evolutiva de esta especie como también
plant polyploids. Bioessays 28: 240–252.
fuera propuesto para otras especies del género DI RIENZO, J. A., F. CASANOVES, M. G.
(Solís Neffa & Fernández, 2001; Elías et al., 2011).
BALZARINI, L. GONZÁLEZ, M. TABLADA &
C. W. ROBLEDO. 2014. iNFOsTAT VERSION
2014. Grupo InfoStat, FCA, Universidad Nacional
de Córdoba, Argentina. http://www.infostat.com.ar.
DURAND, D. & R. HOBERMAN. 2006. Diagnosing
duplications—can it be done? Trends Genet. 22:
Este trabajo fue financiado por proyectos
de la Secretaría General de Ciencia y Técnica ERHENDORFER, F. 1980. Polyploidy and distribution.
(UNNE, PI-17 F189). E. M. S. Moreno es becaria
In: LEWIS, W.H., ed. Polyploidy, biological
Postdoctoral de CONICET-UNNE. V. G. Solís
relevance. New York: Plenum Press, 45–60.
Neffa es miembro de la Carrera del Investigador ELÍAS, G., M. SARTOR & V. G. SOLÍS NEFFA. 2011.
Científico de CONICET.
Patterns of cytotype variation of Turnera sidoides
Bol. Soc. Argent. Bot. 51 (1) 2016
subsp. pinnatifida (Turneraceae) in mountain ranges
Physical Structure, Behaviour and Evolution of
of central Argentina. J. Plant Res. 124: 25-34.
Plant Genomes. Springer; Wien. pp. 255–276.
ELÍAS G., M. SARTOR & V. G. SOLÍS NEFFA. 2011.
INFOSTAT. 2008. InfoStat, version 2008. Grupo InfoStat,
Patterns of cytotype variation of Turnera sidoides
FCA. National University of Córdoba, Argentina.
subsp. pinnatifida (Turneraceae) in mountain ranges KESSLER, M., A. KÜHN, V. G. SOLÍS NEFFA & I.
of central Argentina. J. Plant Res. 124: 25–34.
HENSEN. 2014. Complex geographical distribution
FAVARGER, C. 1967. Cytologie et distribution des
of ploidy levels in Polylepis australis (Rosaceae), an
plantes. Biol. Rev. 42: 163–206.
endemic tree line species in Argentina. Int. J. Plant
FERNÁNDEZ, A. 1973. El ácido láctico como fijador
Sci. DOI: 10.1086/677649.
cromosómico. Bol. Soc. Argent. Bot. 15: 287-290.
KILLEEN, T. J., E. GARCÍA & S. G. BECK. 1993. Guía
FERNÁNDEZ, A. 1987. Estudios cromosómicos en
de árboles de Bolivia. Herbario Nacional de Bolivia
Turnera y Piriqueta (Turneraceae). Bonplandia 6:
& Missouri Botanical Garden.
KÖHLER, C., O. MITTELSTEN SCHEID & A.
FERNÁNDEZ, A. & M. M. ARBO. 1990. Gametas no
ERILOVA. 2011. The impact of the triploid block on
reducidas y relaciones genómicas en tres especies de
the origin and evolution of polyploid plants. Trends
Turnera (Turneraceae). Darwiniana 30: 21-26.
in Genetics 26: 142-148.
FERNÁNDEZ, A. & V. G. SOLÍS NEFFA. 2004. Genomic
KOVALSKY, I. E. & V. G. SOLÍS NEFFA. 2012.
relationships between Turnera krapovickasii (2x, 4x)
Evidence of 2n microspore production in a natural
and Turnera ulmifolia (6x) (Turneraceae, Turnera).
diploid population of Turnera sidoides subsp.
Caryologia 57: 45-51.
carnea and its relevance in the evolution of the T.
FERNÁNDEZ, A., H. REY & V. G. SOLÍS NEFFA.
sidoides (Turneraceae) autopolyploid complex. J. Pl.
2010. Evolutionary realtionships between Turnera
Res. 125: 725–734.
grandiflora and the octoploid T. fernandezii LAZAROFF, Y., A. FERNÁNDEZ & V. G. SOLÍS
(Turneraceae). Ann. Bot. Fen. 47: 321-329.
NEFFA. 2009. Estudios cromosómicos en Turnera
GENTRY, A. H. 1982. Neotropical floristic diversity:
krapovickasii (Turneraceae). Bol. Soc. Argent. Bot.
phytogeographical connections between Central and
South America, Pleistocene climatic fluctuations, or
LAZAROFF, Y., A. FERNÁNDEZ & V. G. SOLÍS
an accident of the Andean orogeny? Ann. Missouri
NEFFA. 2011. Análisis de la producción y frecuencia
Bot. Gard. 69: 557-593.
de gametos no reducidos en poblaciones diploides
GENTRY, A. H. & C. H. DODSON. 1987. Diversity and
de Turnera krapovickasii. J. Basic Appl. Genet.
biogeography of Neotropical vascular epiphytes.
Volumen XLI.
Ann. Missouri Bot. Gard. 74: 205-233.
LEVIN, D. A. 1983. Polyploidy and novelty in flowering
GRANT, V. 1989. Especiación vegetal. Ed. Limusa.
plants. Am. Nat. 122: 1–25.
Méjico, pp. 587.
LEVIN, D. A. 2002. The role of chromosome change in
HAGERUP, O. 1932. Ueber polyploidie in beziehung
plant evolution. New York: Oxford University Press.
zu klima, oekologie und phylogenie. Hereditas 16:
LEWIS, W. H. 1980. Polyploidy in species population. In:
LEWIS, W.H., ed. Polyploidy, biological relevante,
HIJMANS, R. J., L. GUARINO, C. BUSSINK, P.
New York: Plenum Press, 103-144.
MATHUR, M. CRUZ, I. BARRANTES & E.
LÓPEZ, A., A. FERNÁNDEZ, & J. S. SHORE. 2013.
ROJAS. 2004. Diva-Gis. Sistema de Información
Inferences on the origins of polyploid Turnera
Geográfica para el Análisis de Datos de Distribución
species (Passifloraceae) based on molecular data.
Botany 91: 167–175.
HIJMANS, R. J., S. E. CAMERON, J.L. PARRA, P.G.
LUMARET, R., J. L. GUILLERM, J. DELAY, A. AIT
JONES & A. JARVIS. 2005. Very high resolution
IHAJ LOUTFI, J. IZCO & M. JAY. 1987. Polyploidy
interpolated climate surfaces for global landa reas.
and habitat differentiation in Dactylis glomerata L.
Int. J. Climatol. 25: 1965-1978.
from Galicia (Spain). Oecologia 73: 436-446.
HUSBAND, B. C. & D. W. SCHEMSKE. 1998.
MADLUNG, A. 2013. Polyploidy and its effect on
Cytotype distribution at a diploid-tetraploid contact
evolutionary success: old questions revisited with
zone in Chamerion (Epilobium) angustifolium
new tools. Heredity 110: 99–104.
(Onagraceae). Am. J. Bot. 85: 1688-1694.
MEYERS L. A. & D. A. LEVIN. 2006. On the abundance
HUSBAND, B. C., S. J. BALDWIN & J. SUDA.
of polyploids in flowering plants. Evolution 60:
2013. The Incidence of polyploidy in natural
plant populations: Major patterns and evolutionary
MORAES, M. & S. G. BECK. 1992. Diversidad floristica
processes. En: Leitch I. J., J. Greilhuber, J. Doležel,
de Bolivia. En: M. Marconi (ed.). Conservación de la
J. F. Wendel, editors. Plant Genome Diversity 2:
Diversidad Biològica en Bolivia, p. 73-111. Centro
Bol. Soc. Argent. Bot. 51 (1) 2016
Y. Lazzaroff et al. - Análisis citogeográfico en Turnera krapovickasii
de datos para la conservación, La Paz (Bolivia).
SOLÍS NEFFA, V. G. 1996. Cariotipos en Turnera
MORRONE, J. J. 2001. Biogeografía de América Latina
(Turneraceae). Bonplandia 9 (1-2): 121-127.
y el Caribe. M&T-Manuales y Tesis SEA, Zaragoza.
SOLÍS NEFFA, V. G. & A. FERNÁNDEZ. 1993.
NAVARRO, G. & M. MALDONADO. 2002. Geografía
Estudios cromosómicos en especies de Turnera
Ecológica de Bolivia: Vegetación y Ambientes
(Turneraceae). Bonplandia 7 (1-4): 101-118.
Acuátcos. Editorial: Centro de Ecología Simón I.
SOLÍS NEFFA, V. G. & A. FERNÁNDEZ. 2000.
Patiño-Departamento de Difusión. Cochabamba,
Chromosome studies in Turnera (Turneraceae).
Genet. Mol. Biol. 23: 925-930.
OSBORN, T. C., D. V. BUTRULLE, A. G. SHARPE,
SOLIS NEFFA, V. G. & A. FERNÁNDEZ. 2001.
K. J. PICKERING, I. A. PARKIN, J. S. PARKER
Cytogeography of the Turnera sidoides L. complex
& D. J. LYDIATE. 2003. Detection and effects of
(Turneraceae, Leiocarpae). Bot. J. Linn. Soc .137:
a homeologous reciprocal transposition in Brassica
napus. Genetics 165: 1569–1577.
SOLÍS NEFFA, V. G., A. F. PANSERI, W. REYNOSO
PANSERI, A. F., J. G. SEIJO & V. G. SOLÍS NEFFA.
& J. G. SEIJO. 2004. Variación en el color de flores
2008. Análisis de la producción y frecuencia de
y números cromosómicos en el Noroeste del área
microsporas no reducidas en diploides de Turnera
de distribución de Turnera sidoides (Turneraceae).
sidoides (Turneraceae). Bol. Soc. Argent. Bot. 43:
Bonplandia 13: 117–128.
SOLTIS, D. E. 1984. Autopolyploidy in Tolmiea menziesii
RAMAN, V. S. & P. C. KESAVAN. 1964. Meiosis and
(Saxifragaceae). Am. J. Bot. 71: 1171–1174.
nature of polyploidy in Turnera ulmifolia. J. Indian SOLTIS, D. E., P. S. SOLTIS & J. A. TATE. 2004.
Bot. Soc. 43: 495-499.
Advances in the study of polyploidy since plant
RAMELLA, L. & R. SPICHIGER. 1989. Interpretación
speciation. New Phytol. 161: 173–191.
preliminar del medio físico y de la vegetación del STEBBINS, G. L. 1971. Chromosomal evolution in
Chaco Boreal. Contribución al estudio de la flora y
higher plants (London: Addison-Wesley Press).
de la vegetación del Chaco. I. Candollea 44: 639-
TISCHLER, G. 1935. Die bedeutung der polyploidie
für die verbreitung der Angiospermen, erläutert an
RAMSEY, J. & D. W. SCHEMSKE. 1998. Pathways,
den arten Schleswig-Holsteins, mit Ausblicken auf
mechanisms, and rates of polyploid formation in
andere florengebiete. Bot. Jahrb. Syst. 67: 1–36.
flowering plants. Annu. Rev. Ecol. System. 29: 467-
UDALL, J. A. & J. F. WENDEL. 2006. Polyploidy and
crop improvement. Crop Sci. 46: S3-S14.
REYNOSO, W. L., V. SOLÍS NEFFA & A. FERNÁNDEZ.
URBAN, I. 1883. Monographie der familie der
2004. "Análisis preliminar de la distribución
Turneraceen. Jahrb. Königl. Bot. Gart. Berlin 2:
geográfica de los citotipos de Turnera krapovickasii
(Turneraceae)". VIII Comunicaciones Científicas y
VAN DIJK, P., M. HARTOG & W. VAN DELDEN. 1992.
Tecnológicas de la UNNE.
Single cytotype areas in autopolyploid Plantago
RIBERA ARISMENDI, M. 1993. Regiones ecológicas.
media L. Biol. J. Linn. Soc. 46: 315–331.
En: M. Marconi (ed.) Conservación de la Diversidad
WENDEL, J. & J. DOYLE. 2005. Polyploidy and
Biológica en Bolivia, p. 9-71. Centro de datos para la
evolution in plants. P. 97–117. En R.J. Henry
conservación, La Paz (Bolivia).
(ed.) Plant diversity and evolution: Genotypic
SHORE, J. S. 1991a. Chromosomal evidence for
and phenotypic variation in higher plants. CAB
autotetraploidy in the Turnera ulmifolia complex
(Turneraceae). Can. J. Bot. 69: 1302-1308.
YOUNG, K. E.; C. ULLOA ULLOA, J. L. LUTEYN &
SHORE, J. S. 1991b. Tetrasomic inheritance and isozyme
S. KNAPP. 2002. Plant evolution and endemism in
variation in T. ulmifolia vars. elegans Urb. and
Andean South America: an introduction. Bot. Rev.
intermedia Urb. (Turneraceae). Heredity 66: 305-
SHORE, J. S. & S. C. H. BARRET. 1986. Genetic
modification of dimorphic incompatibility in the
Turnera ulmifolia L. complex (Turneraceae). Canad. Recibido el 24 de julio de 2015, aceptado el 23 de
J. Genet. Cytol. 28: 796 - 807.
septiembre de 2015.
Source: http://www.botanicargentina.com.ar/boletin/511/13_lazzaroff.pdf
ORIGINAL CONTRIBUTION Adjunctive Risperidone Treatmentfor Antidepressant-Resistant Symptomsof Chronic Military Service–Related PTSDA Randomized Trial John H. Krystal, MD Context Serotonin reuptake-inhibiting (SRI) antidepressants are the only FDA- Robert A. Rosenheck, MD approved pharmacotherapies for the treatment of posttraumatic stress disorder (PTSD).
PTH/PTHrP Receptor Mediates Cachexia in Modelsof Kidney Failure and Cancer Graphical Abstract Serkan Kir, Hirotaka Komaba,Ana P. Garcia, ., Beate Lanske,Richard A. Hodin, Bruce M. Spiegelman In BriefKir et al. reveal a role for the PTH/PTHrPpathway in cachexia driven by kidneyfailure or cancer and show how PTH andPTHrP stimulate adipose tissue browningthrough their receptor PTHR. Loss ofPTHR in adipocytes blocks atrophy of notonly fat tissue, but also skeletal muscle inmice.